What the economy is to human society, metabolism is to the cell. In short, metabolism is the ensemble of chemical processes by which cells obtain the goods and services required for their continuance, growth and reproduction.

Franklin M. Harold
The Way of the Cell (p. 73). Oxford University Press. Kindle Edition.
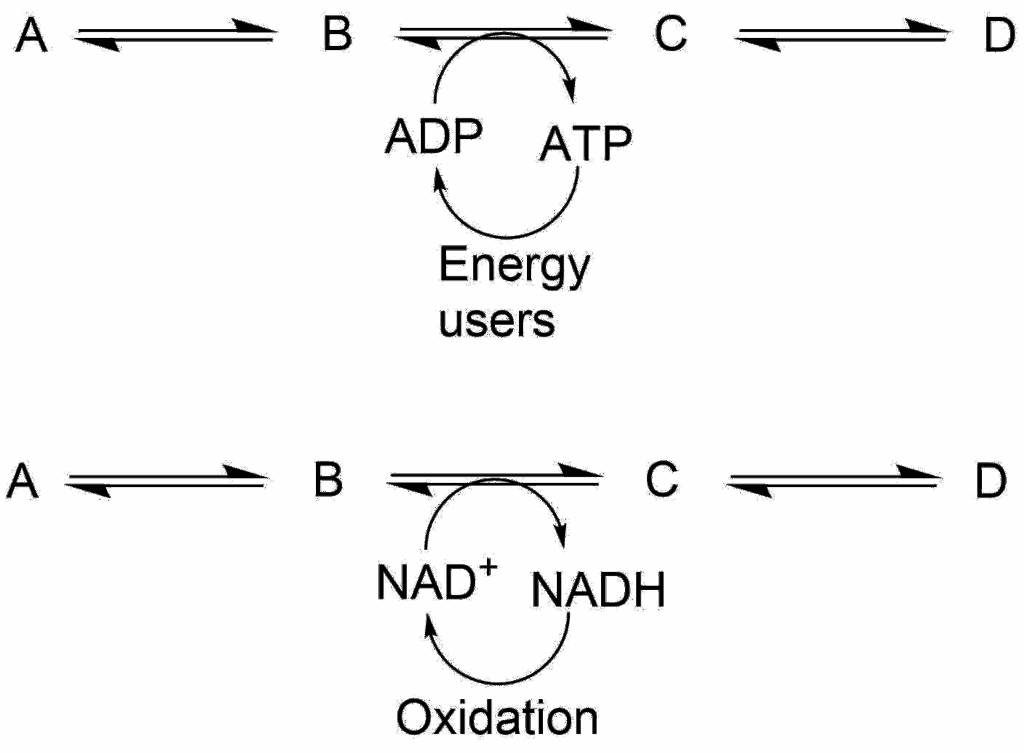
As in economics, the linkage between supply and demand is intensely dynamic. The great highways of energetics, respiration and photosynthesis, keep the ATP/ADP ratio high, far away from equilibrium, and that, in turn, allows ATP to serve as an energy donor, displacing from equilibrium those reactions in which it participates. All biosynthetic processes, and also those that entail movement or transport, are energized either by ATP or by one of the more specialized energy carriers, and the latter are linked to the ATP/ADP couple, as it were by a system of exchange rates.
Harold, Franklin M.
The Way of the Cell (p. 42). Oxford University Press. Kindle Edition.
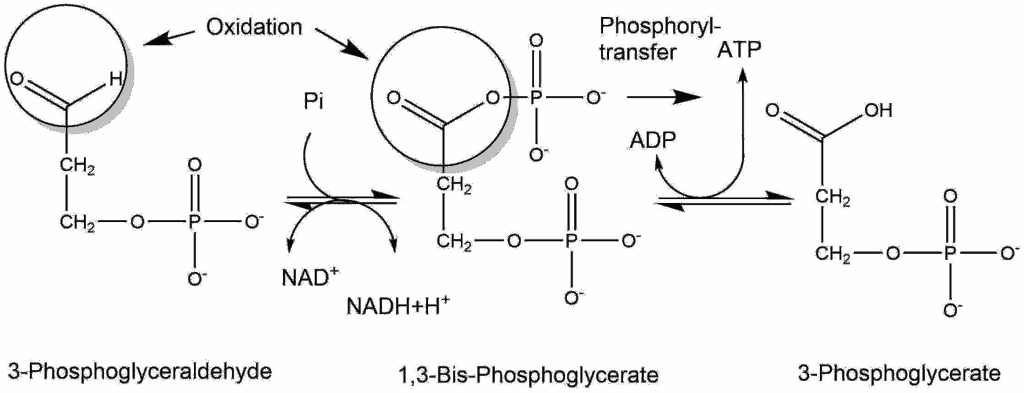
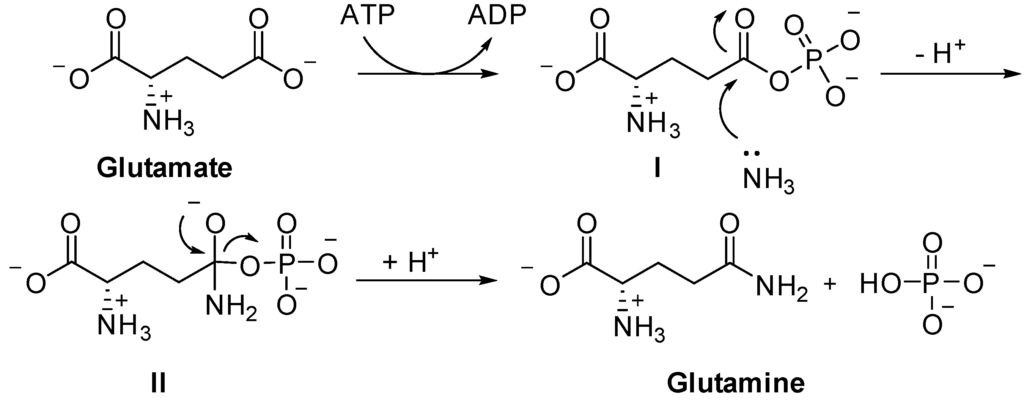
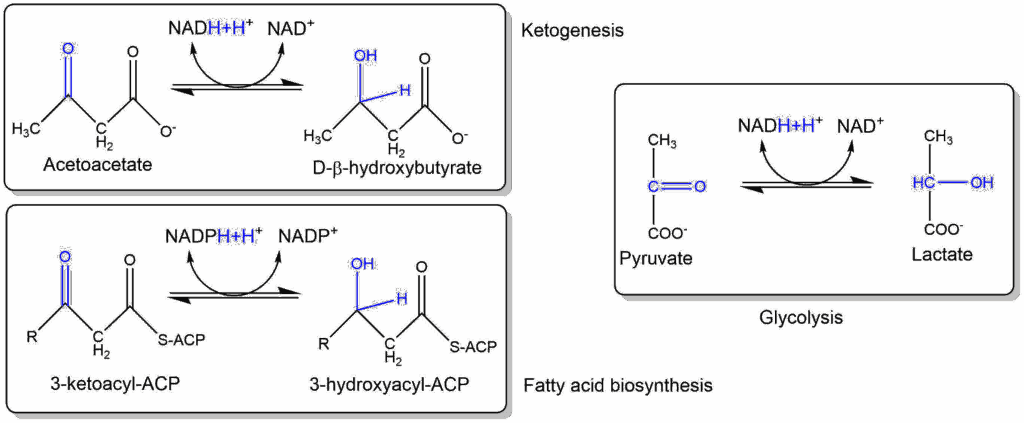
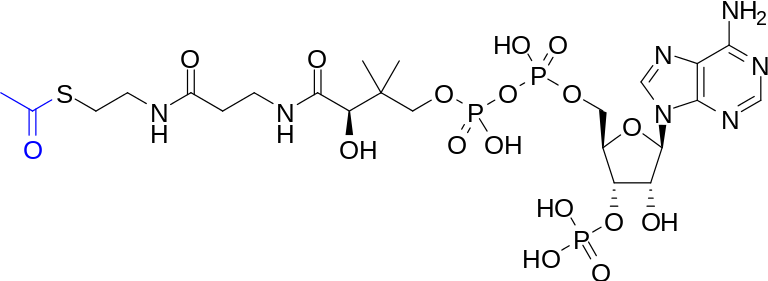
It is revealing that thioesters are obligatory intermediates in several key processes in which ATP is either used or regenerated. Thioesters are involved in the synthesis of all esters, including those found in complex lipids. They also participate in the synthesis of a number of other cellular components, including peptides, fatty acids, sterols, terpenes, porphyrins, and others. In addition, thioesters are formed as key intermediates in several particularly ancient processes that result in the assembly of ATP. In both these instances, the thioester is closer than ATP to the process that uses or yields energy. In other words, thioesters could have actually played the role of ATP in a "thioester world" initially devoid of ATP. Eventually, [these] thioesters could have served to usher in ATP through its ability to support the formation of bonds between phosphate groups.

Christian de Duve
(1995). "The Beginnings of Life on Earth". American Scientist. 83(5): 428–437. Bibcode:1995AmSci..83..428M.
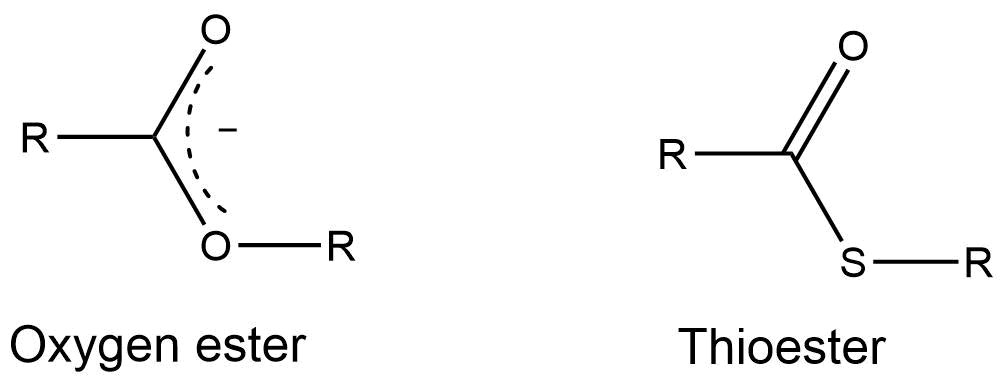
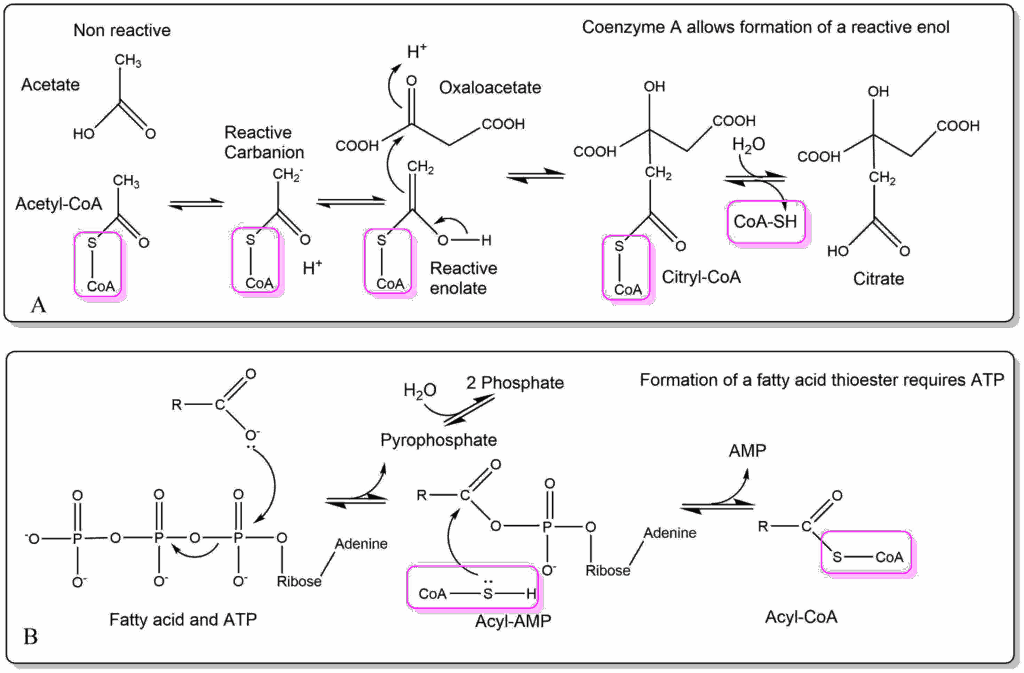
Question
Answer
Question
Try to write down the biochemical reaction mechanism that would form an ester between a fatty acyl-CoA molecule and glycerol-3-phosphate (at the positions where hydroxyl groups are).
Answer
![]()
{kind=link}
![User:Bryan Derksen (original) and DMacks (talk) (color-change) [Public domain], via Wikimedia Commons](https://commons.wikimedia.org/wiki/File:Acetyl-CoA-2D_colored.svg){kind=link}