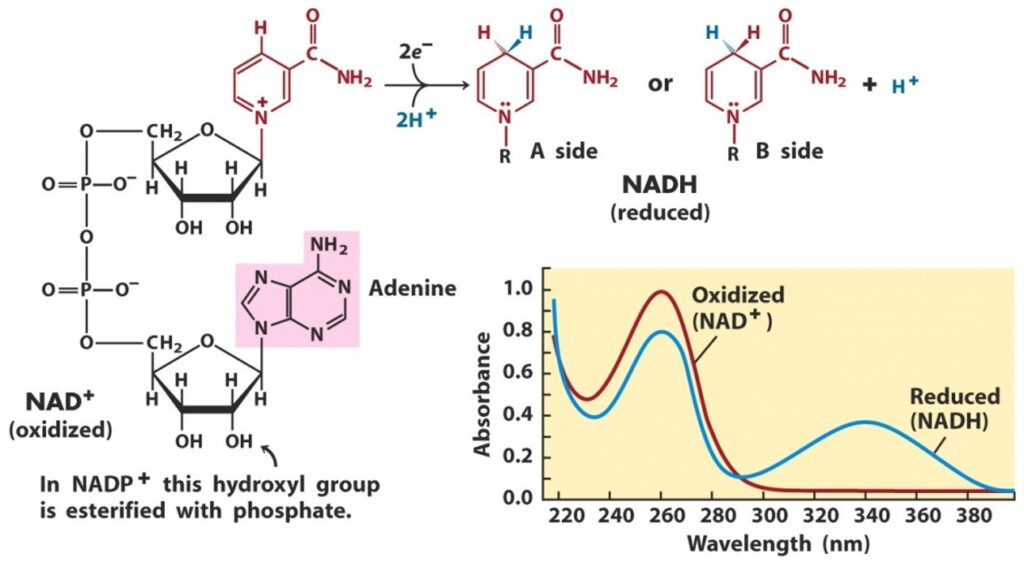
What do you think the time course would look like?
Draw curves by hand on a piece of paper for glucose concentrations of: 10 µM, 100 µM, and 1000 µM.
The curves do not need to be exact but should have the right shape and relative end points.
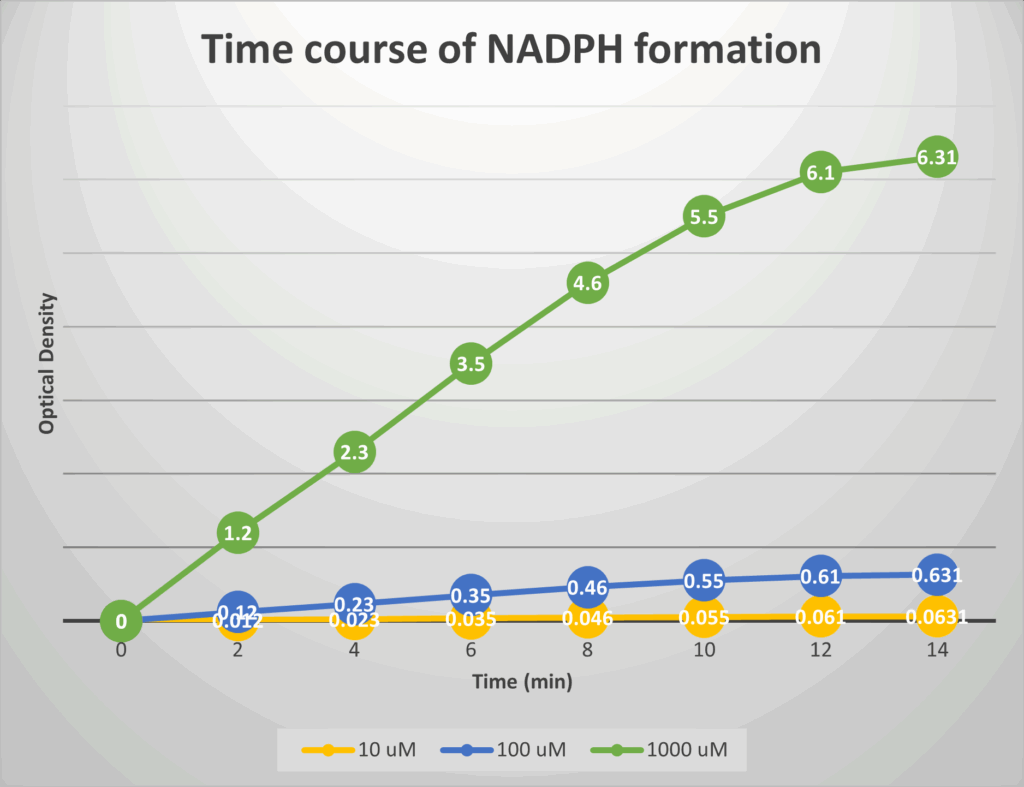



Draw on paper or generate and Excel sheet that plots the reaction rate at concentrations (uM): 0, 20, 50, 100, 200, 500, 1000, 2000, 3500, 5000, 7500 and 10000 for enzyme orange (Km of 2000 uM and Vmax of 100).
Place a vertical line at the position where Enzyme Orange has its Km-value (remember enzyme Orange has the same Vmax as enzyme Blue). What happens to the Km-value when the Vmax of enzyme Orange is increased from 100 to 200 and to 400?
Does a change of Vmax affect the Km-value?
The Km-value does not change.

Start from the kinetic behavior of Enzyme Orange (Vmax = 100, Km = 2000 uM).
What happens to the curve when you set the Km to 500 uM and 10000 uM?
As the Km increases, higher substrate concentrations are needed to reach Vmax.

Allosteric enzymes are multimeric and the subunits influence each other. Increase the number of subunits from 1 to 4 and look at the response of the enzyme to changes of substrate concentration.
Note: The response to changes of the number of subunits is idealised. For instance, the real behaviour of an enzyme with two subunits would look more like 1.5 or 1.6 (which you can test) an enzyme with three subunits would look like 2.5 etc.
How does the curve change?
As the number of subunits increase the sigmoidity of the curve increases.

