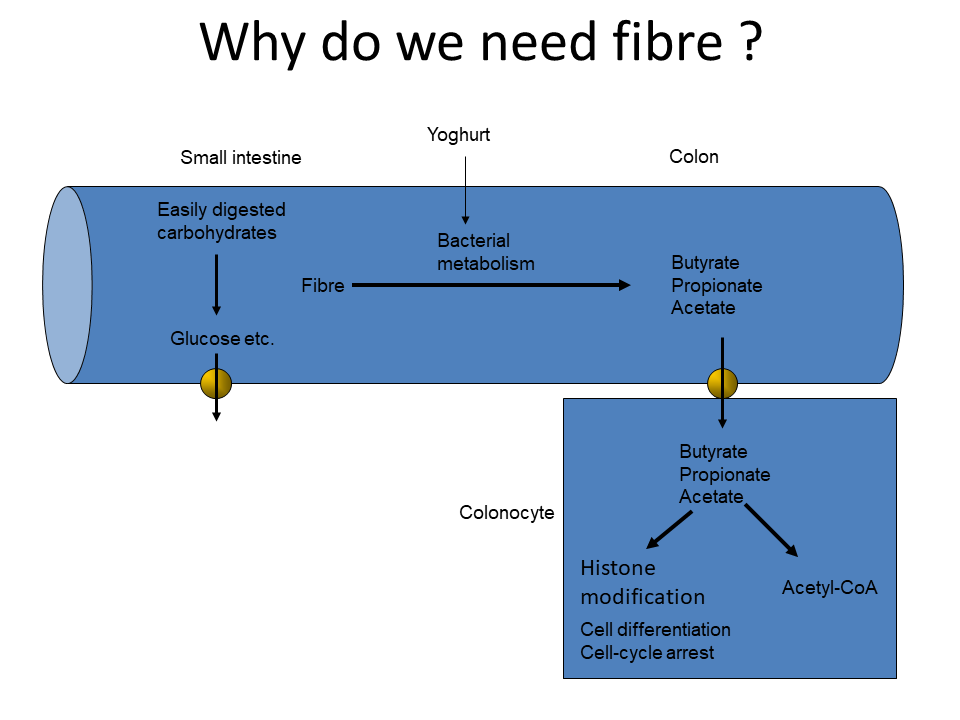
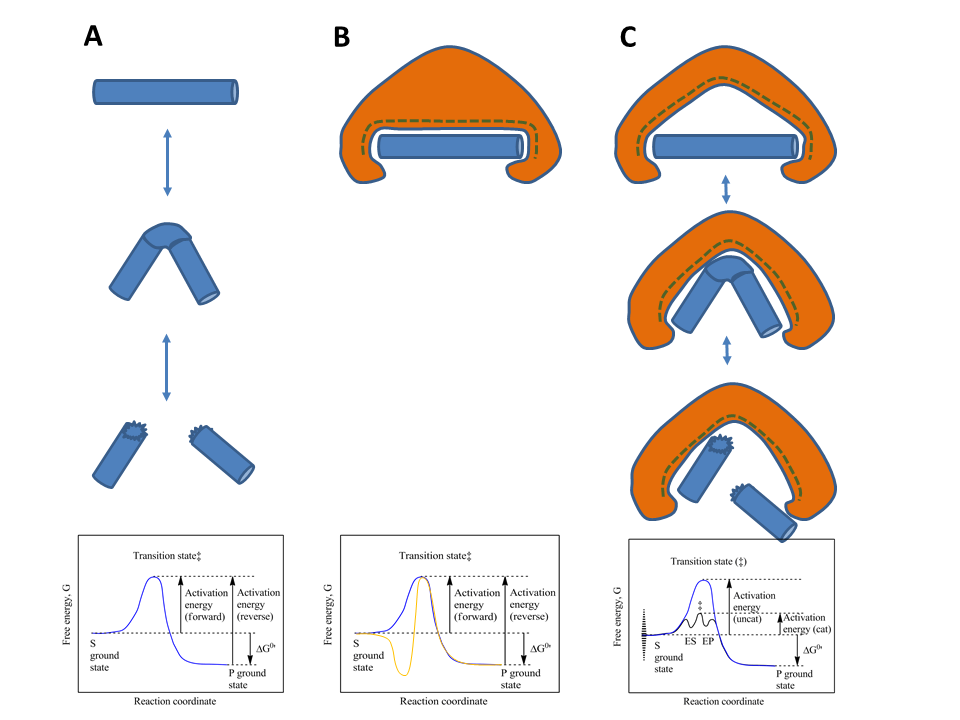
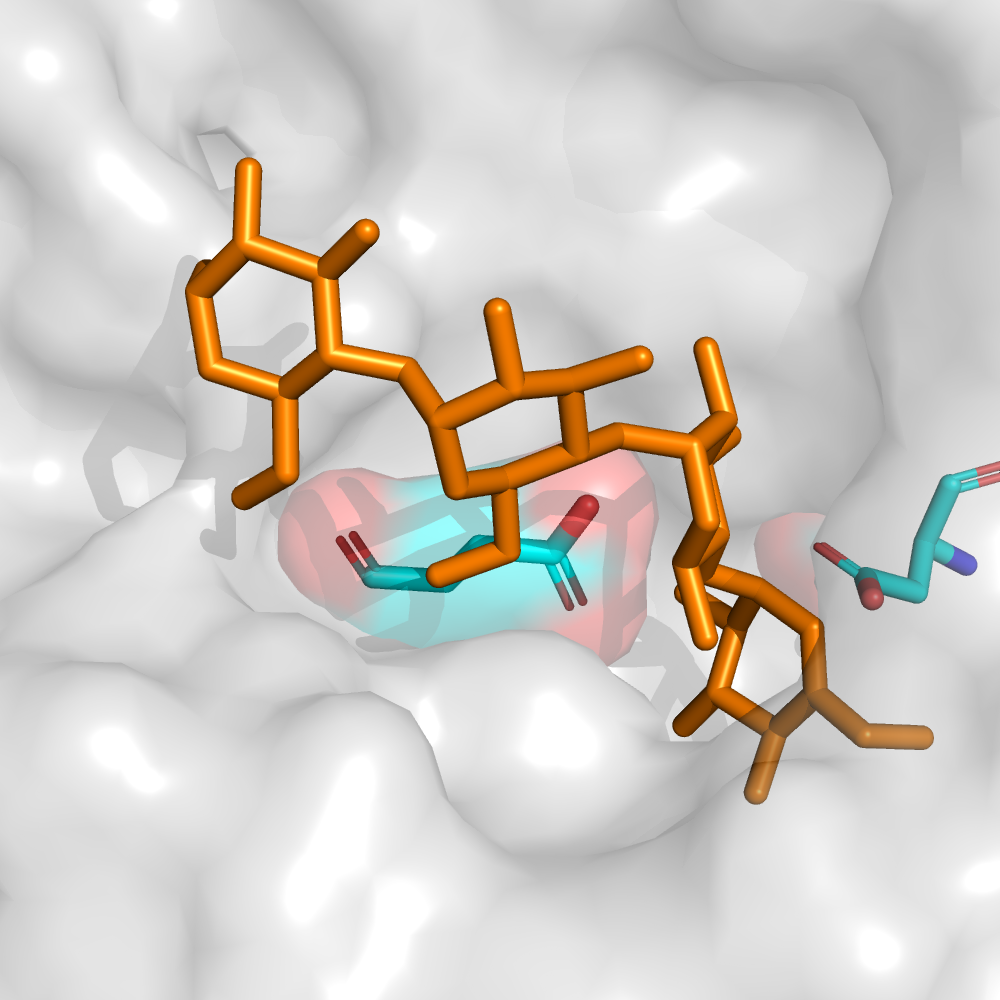
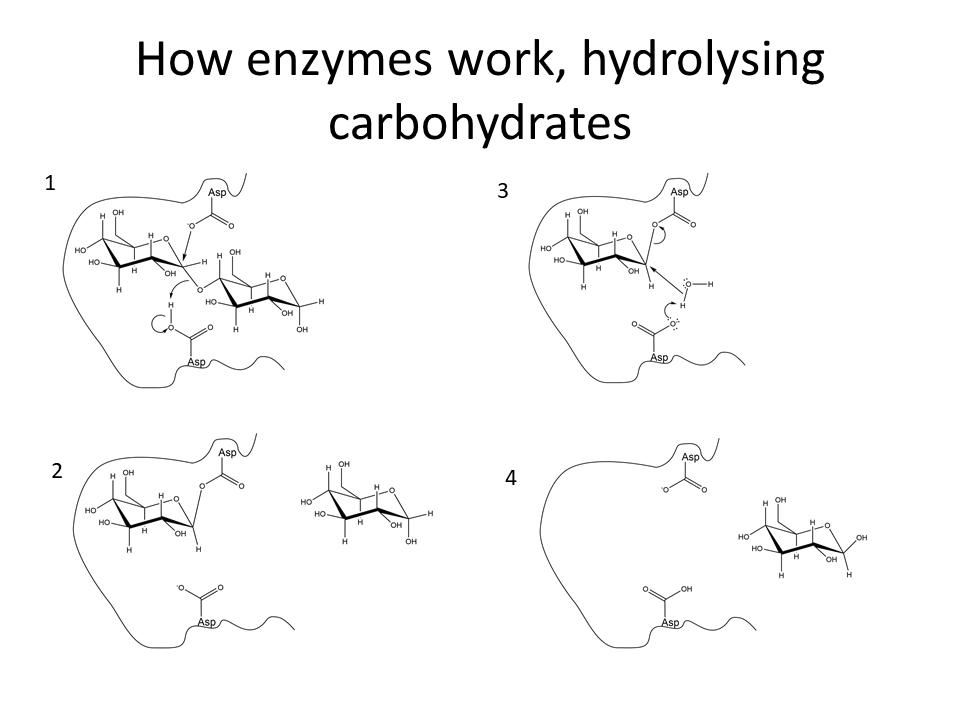
Beans, peas, soybeans and other leguminous plants contain oligosaccharides with (1,6)-linked galactose residues that cannot be hydrolysed by human enzymes. What is the fate of these polysaccharides in the intestine.
These carbohydrates will not be absorbed in the small intestine, but will be used by bacteria in the large intestine. The bacteria will convert the carbohydrates into hydrogen gas, lactic acid and short-chain fatty acids. The gas generated by legume consumption is well-known.