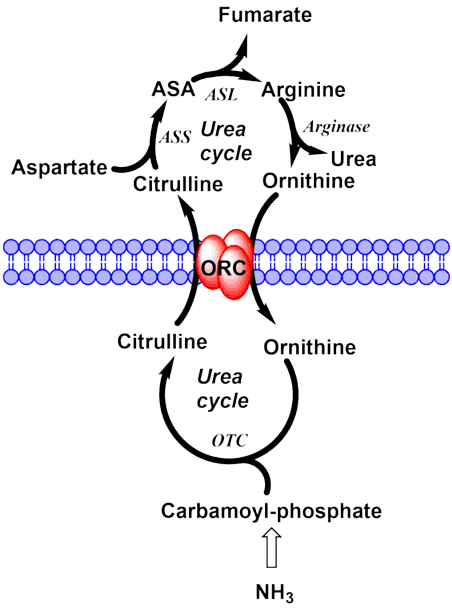
We can analyse transport processes in the same way as enzymatic reactions. We will discuss this formal analysis of enzymatic reactions in the next chapter. The reaction can be written as
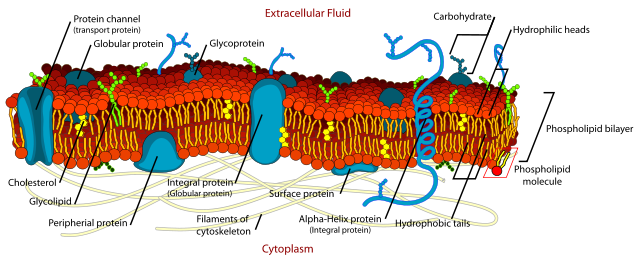
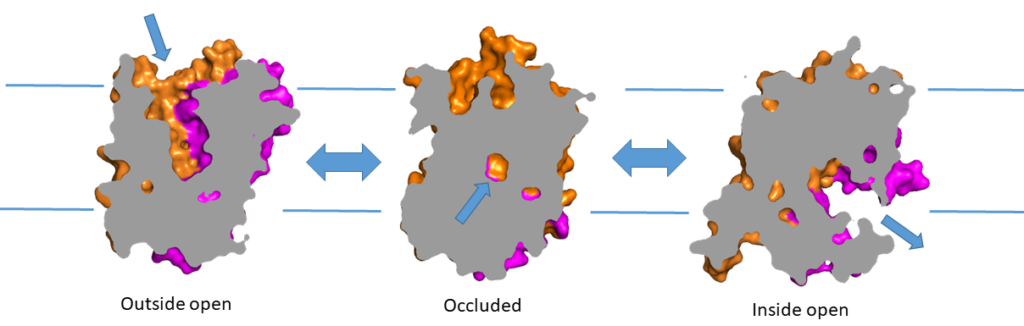
In chapter 4 you discovered the secondary structure elements of soluble proteins. These elements are also found in membrane transporters (Fig. 8 A,B). Alpha-helices are typically used to span the lipid membrane. Because of the hydrophobic environment transmembrane alpha-helices predominantly contain non-polar amino acids. This can be seen in Fig. 8D, in which atoms that generate polarity (oxygen, Red; nitrogen, Blue) are mainly found in the caps of the protein, which are in contact with water. Side chains that are entirely formed by aliphatic amino acids are shown in light-blue color (carbon-hydrogen), sulphur atoms which are less polar than nitrogen and oxygen are shown in yellow. The transmembrane helices are arranged to form a tunnel through which the substrate can reach the binding site (Fig. 8C). Narrow turns connect the membrane-spanning helices. Larger loops provide flexibility to allow conformational changes.