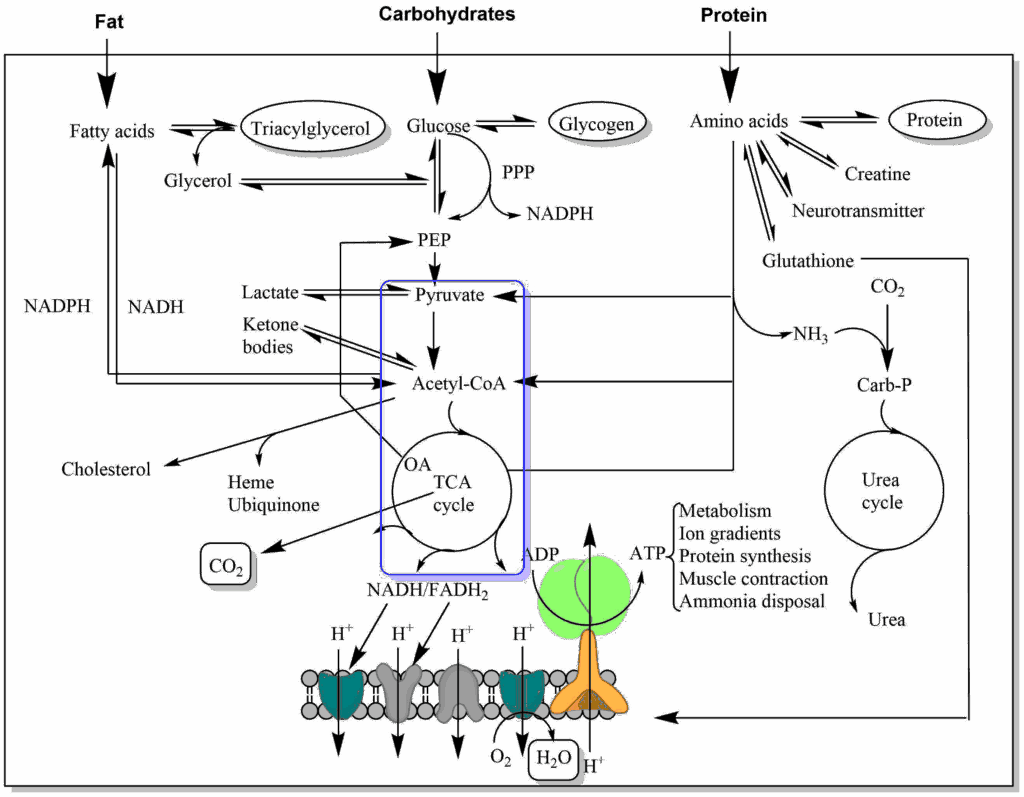
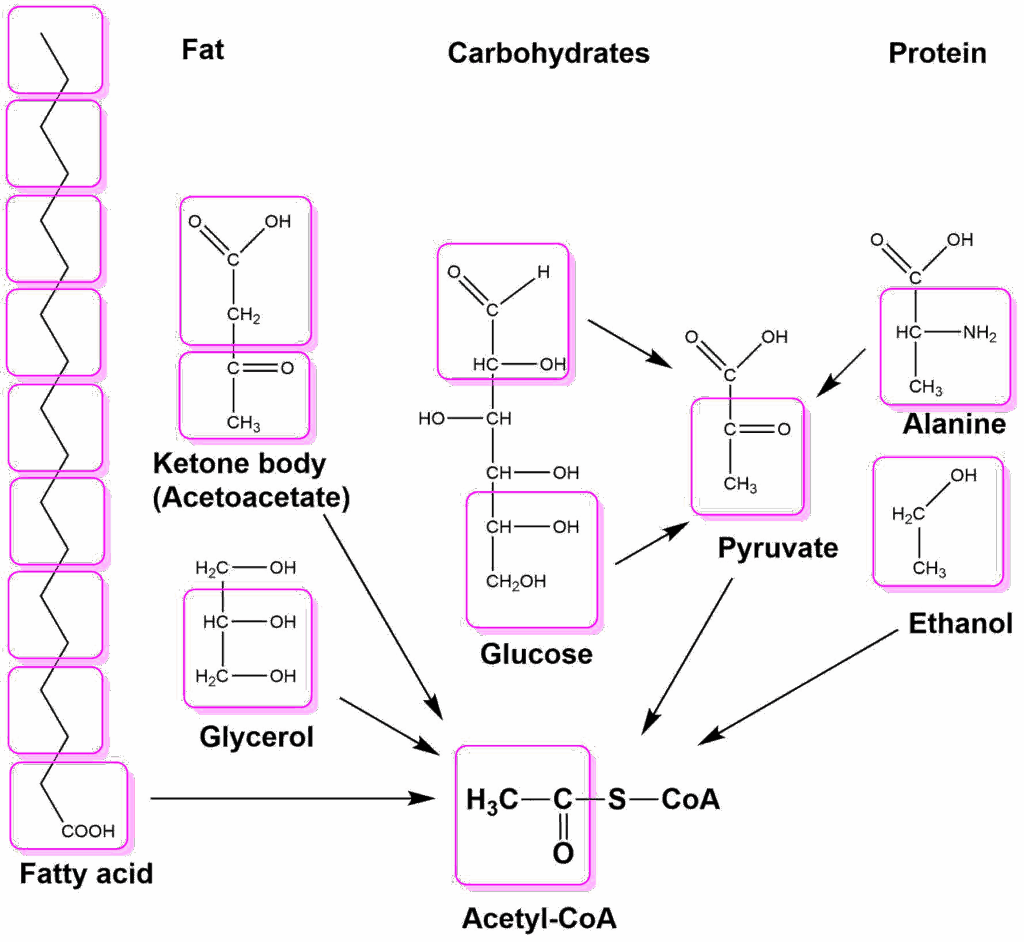
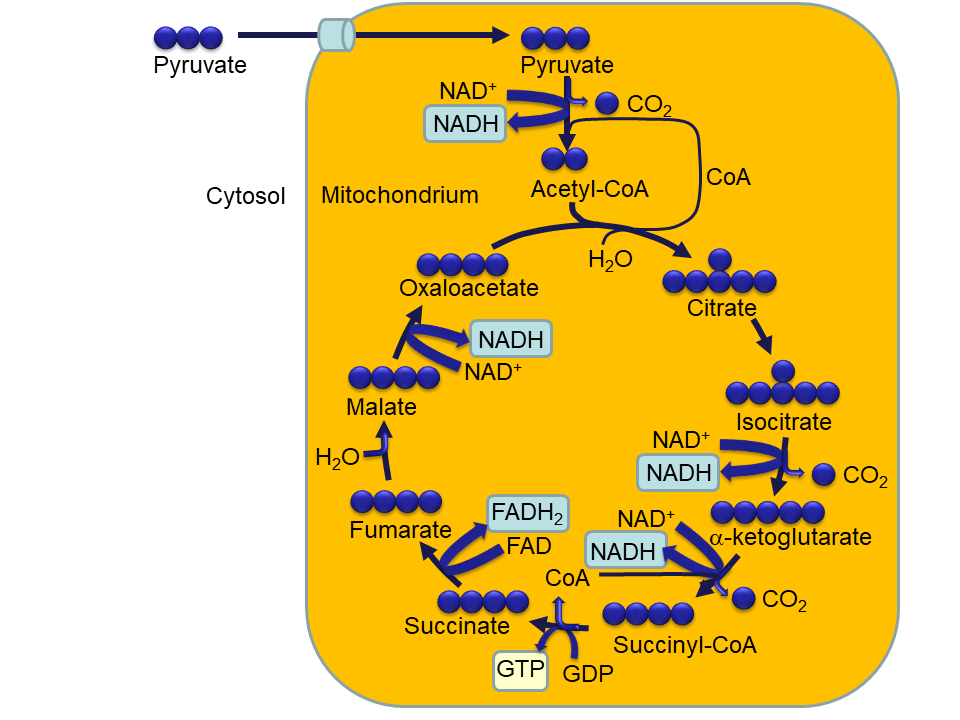
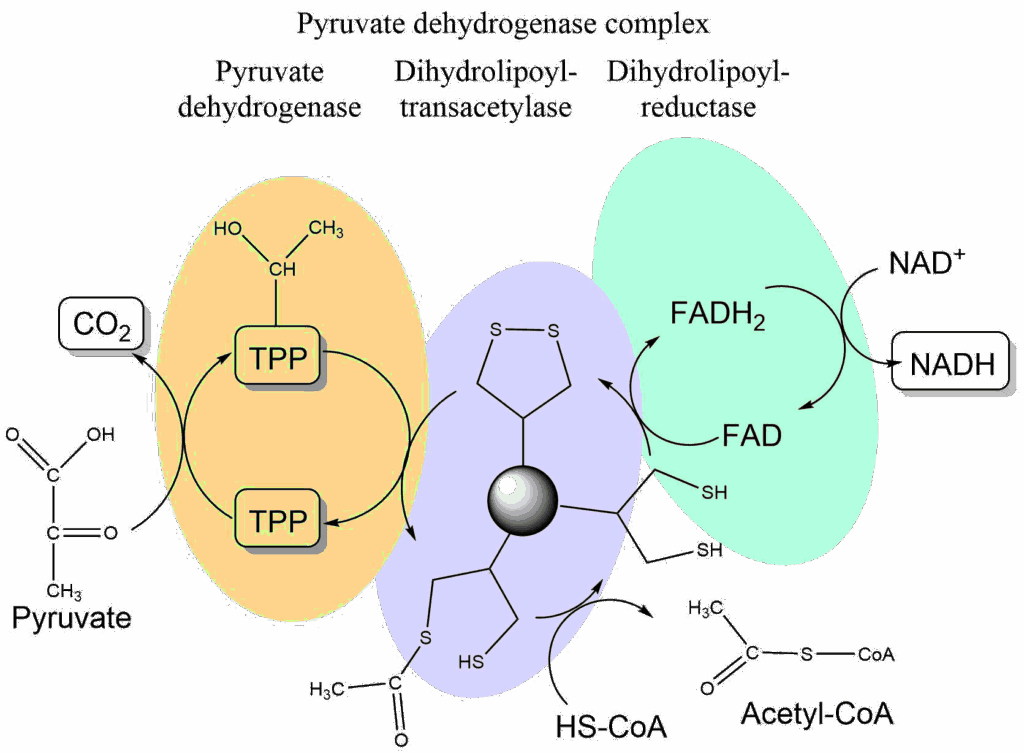
Despite what you might guess, when monitoring your breathing, your body doesn't care whether you're inhaling enough oxygen. It cares only whether you're expelling enough carbon dioxide - that's the gas that sets off the panic button when you're suffocating.

Sam Kean
Author
Question
Answer
Question
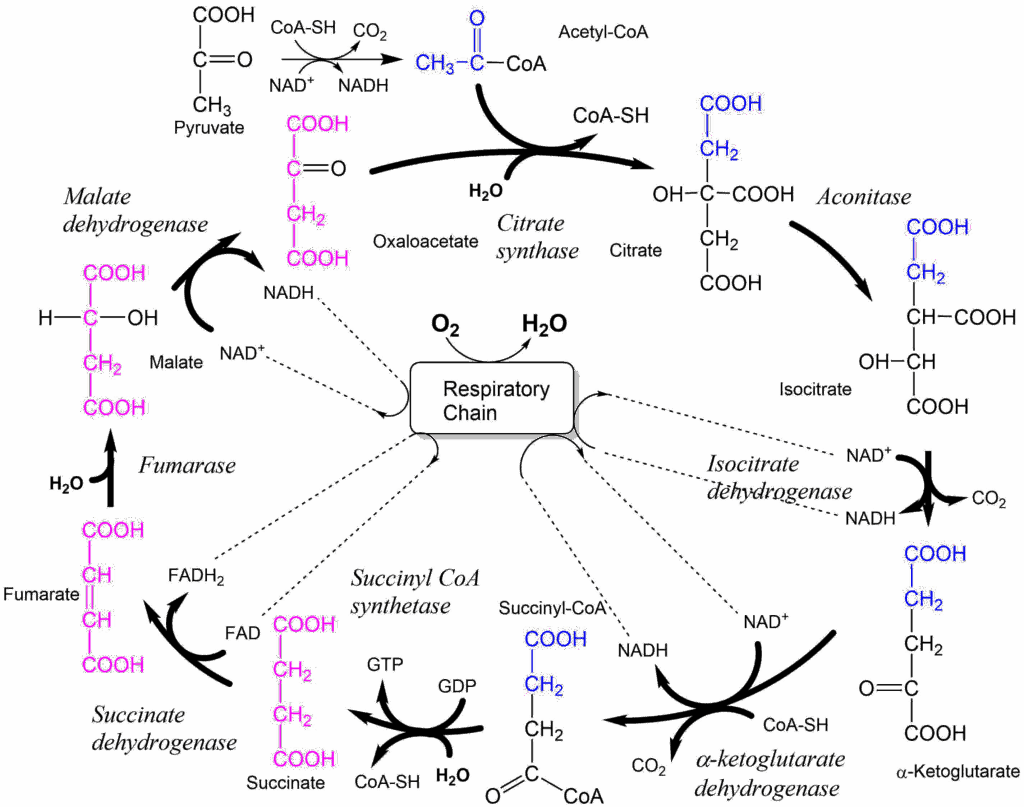
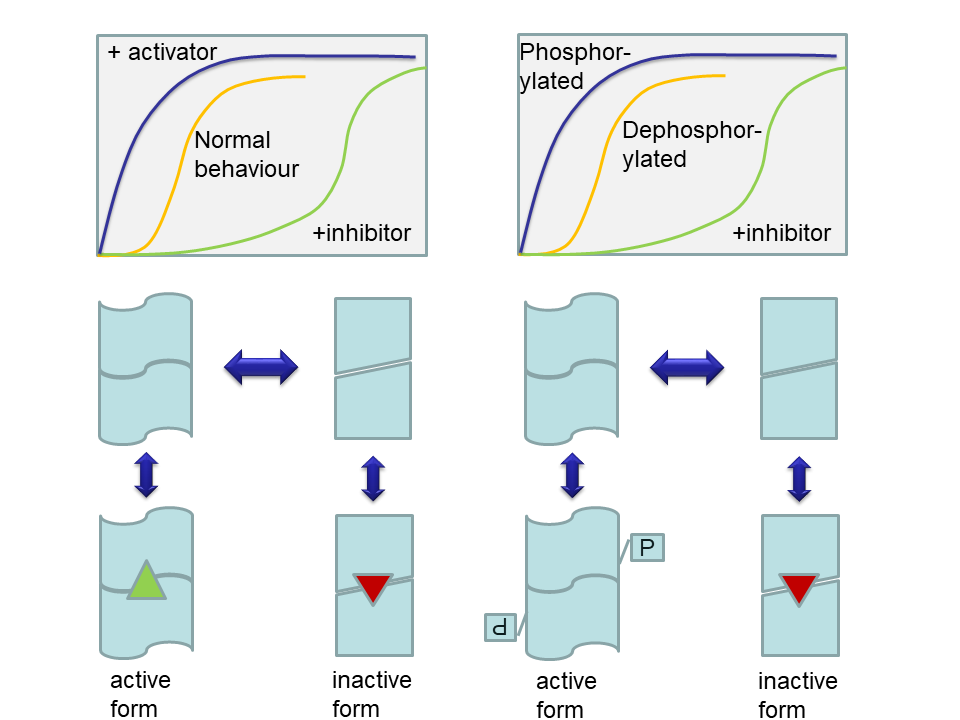
In an experiment you change the concentration of isocitrate to measure the reaction rate of isocitrate dehydrogenase. You get the following graph.
- What is the behaviour of isocitrate dehydrogenase called? How do you recognise the behaviour?
- Draw a graph of enzyme rate vs. ADP concentration at an isocitrate concentration of 0.5 mM.
- Draw a graph of enzyme rate vs. NADH concentration at an isocitrate concentration of 0.5 mM.
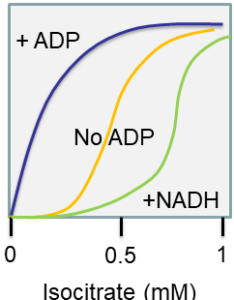
Answer
- Allosteric behaviour. It can be recognised by the sigmoidal shape of the relation between reaction rate and substrate concentration.
- The graph would start at the rate achieved by the enzyme in the absence of ADP (about 0.5 x Vmax) and would increase in a hyperbolic shape to the maximum rate achieved by the stimulated enzyme (Vmax).
- The graph would start at the rate achieved by the enzyme in the absence of NADH (about 0.5 x Vmax) and would decrease exponentially to the minimum rate of the less active enzyme (0.1 x Vmax).
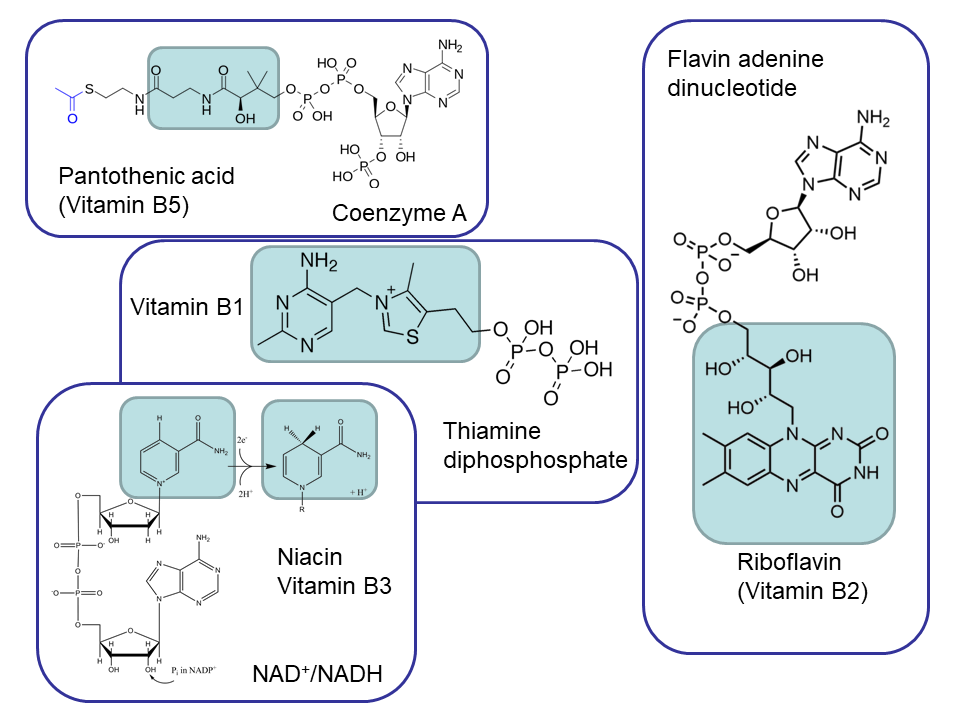
![User:Bryan Derksen (original) and DMacks (talk) (color-change) [Public domain], via Wikimedia Commons.](https://commons.wikimedia.org/wiki/File:Acetyl-CoA-2D_colored.svg){kind=link}
![Edgar181 [Public domain], from Wikimedia Commons](https://commons.wikimedia.org/wiki/File:Thiamine_diphosphate.png){kind=link}
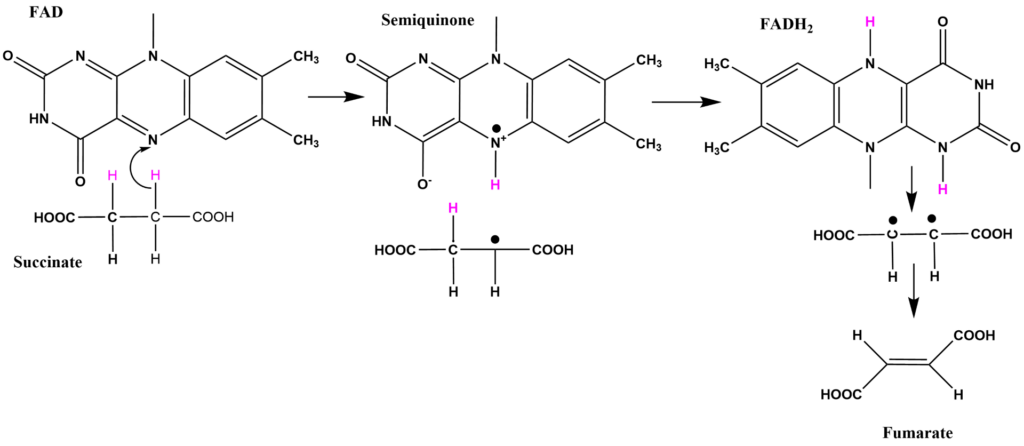
![Edgar181 [Public domain], from Wikimedia Commons](https://commons.wikimedia.org/wiki/File:Flavin_adenine_dinucleotide_H2.png){kind=link}
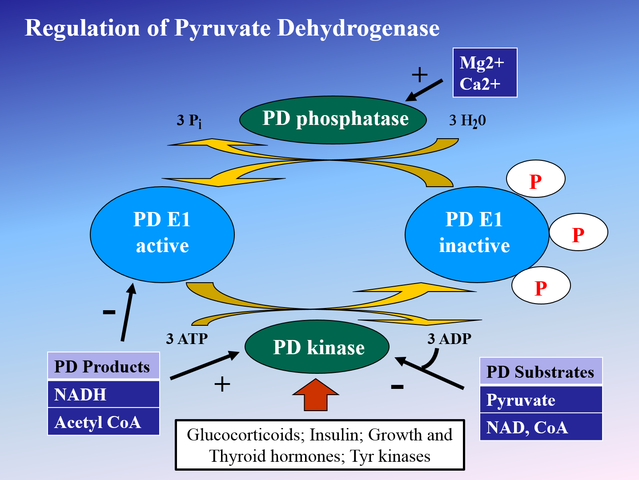
![Rmostell Ray Mosteller [Public domain], from Wikimedia Commons](https://commons.wikimedia.org/wiki/File:1400x1048_pdh_regulation.png){kind=link}