1 Building Blocks of tissues and cells
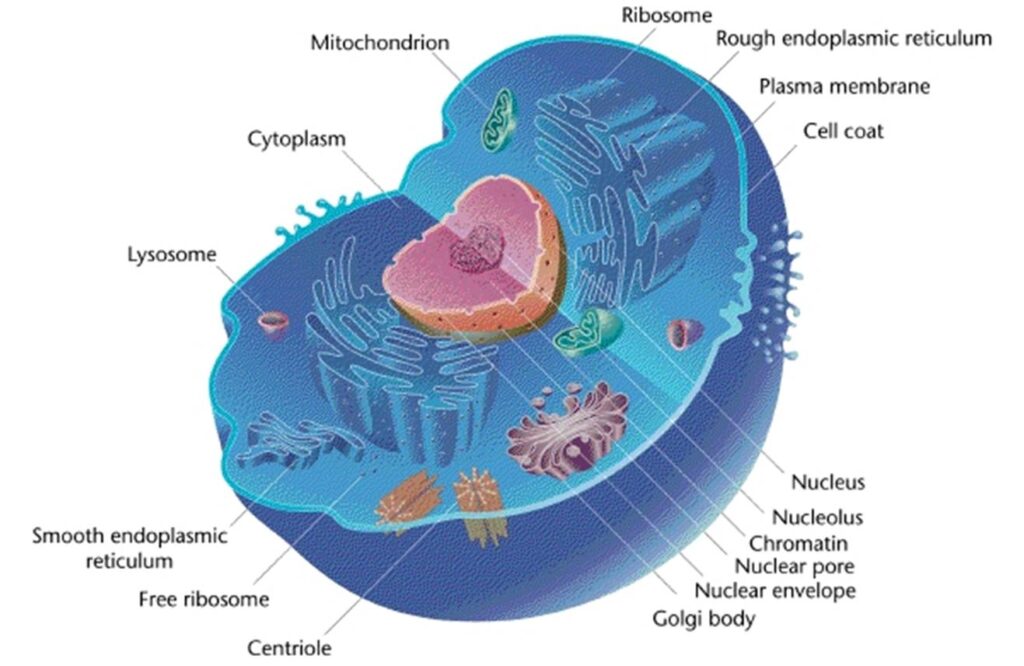
Before we follow the digestion and metabolism of nutrients, we need to discuss what building blocks are required to make a cell. Please note that our food is derived from plant cells or animals cells and therefore has the very same building blocks that are used to make our own cells.
Learning goals
- Understand what is required to make a cell.
- Appreciate the chemical structure of biomolecules.
- Titration of amino acids.
- Get a first impression of the purposes of biomolecules.
Key concepts
- Dynamics of a living cell
- Physico-chemical properties of amino acids
- Physico-chemical properties of lipids
- Building blocks of a cell
Building blocks of tissues and cells
Nutrition is needed to provide energy, but also to provide building blocks that make up tissues and cells. It is an instructive exercise to consider what compounds are required to build an organism. To make a new cell we need (Table 1):
| Building block | Structure |
|---|---|
|
Amino acids |
Protein |
|
Nucleotides |
DNA, RNA |
|
Lipids |
Membranes, Storage |
|
Carbohydrates |
Storage, protein and lipid modification, nucleotides |
|
Vitamins |
Enzyme cofactors, antioxidants |
|
Minerals, Metals |
Enzyme cofactors, modification of molecules and proteins, cytosolic ions, oxygen transport, modification of molecules and proteins |
Watch this video to get an impression of how cells work and how dynamic they are, when looked at a high magnification. You will not understand all the details at this point, but the animation shows an immune cell rolling along the wall of a blood vessel. The observer then moves to the membrane and looks into the cell and its cytoskeleton. Next you will look at the synthesis of protein using mRNA as a template. Then watch how some proteins are inserted into the membrane of the endoplasmic reticulum. The membranes with proteins bud off and move to the plasma membrane where they fuse and release contents.
We have ample reason to believe that every biological phenomenon, however complex, is ultimately based on chemical and physical interactions among molecules. Harold, Franklin M.. The Way of the Cell (p. 4). Oxford University Press. Kindle Edition.
In figure 1 and the video the cell looks quite empty. Is this real. Figure 2 shows an illustration of the crowded environment in a living cell.
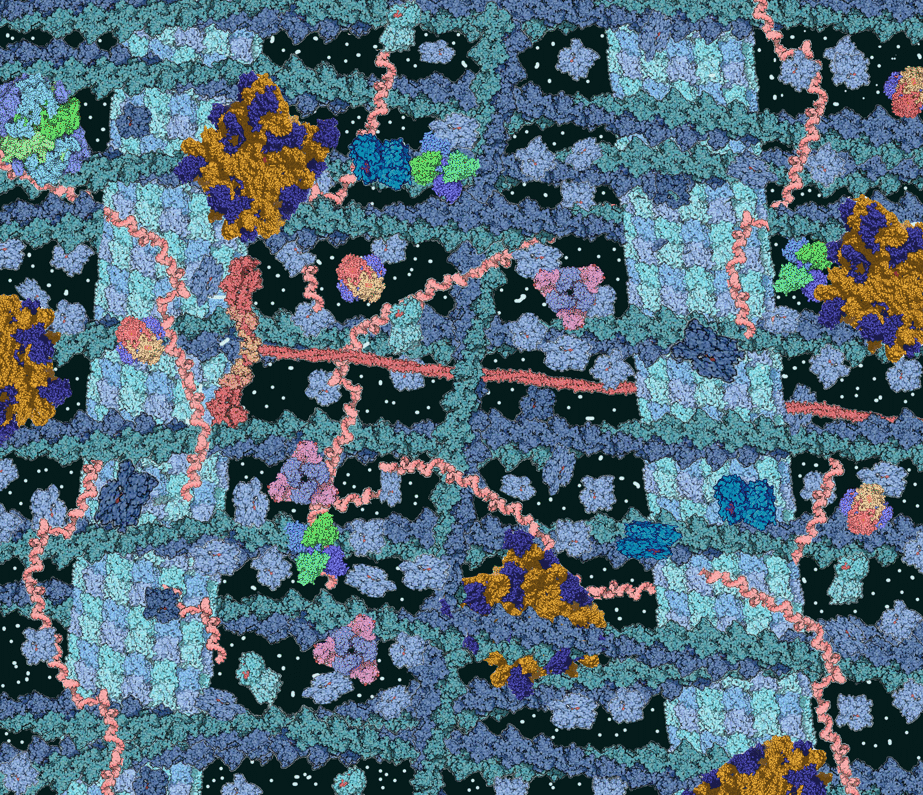
Protein is quantitatively the largest component of any cell. It includes all enzymes to carry out metabolism, signalling proteins, defence proteins, structural proteins and storage proteins. All proteins are made up of 20 different amino acids (Fig. 3)
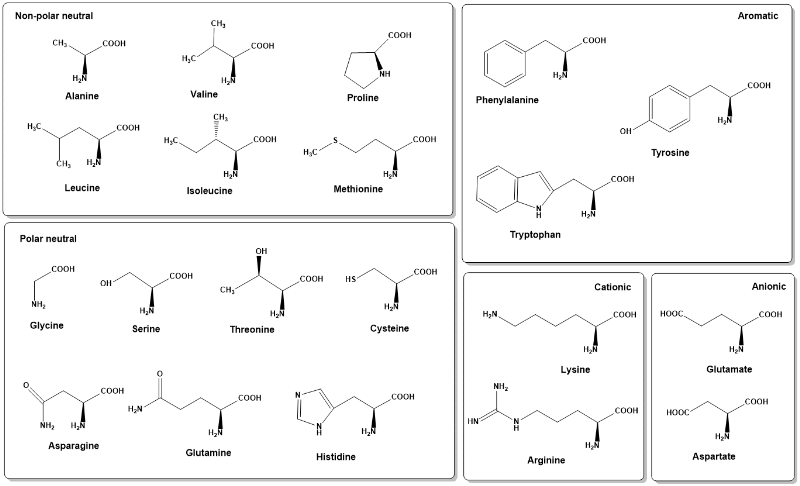
Table 2: Acid-base properties of amino acids
| Functional group | pKa (approximate) | Amino acid |
|---|---|---|
|
α-carboxyl-group |
3.1 |
all |
|
Terminal carboxyl-group |
4.1 |
aspartate, glutamate |
|
Imidazole |
6.0 |
histidine |
|
Sulfhydryl |
8.3 |
cysteine |
|
α-amino group |
9.0 |
all |
|
Phenylhydroxy-group |
10.9 |
tyrosine |
|
Terminal amino-group |
10.5 |
lysine |
|
Guanidinium |
12.5 |
arginine |
Watch the following video to see the titration of the amino acid glycine. In Figure 4 you can then look at the more complex acid/base properties of histidine, which has 3 ionizable groups.
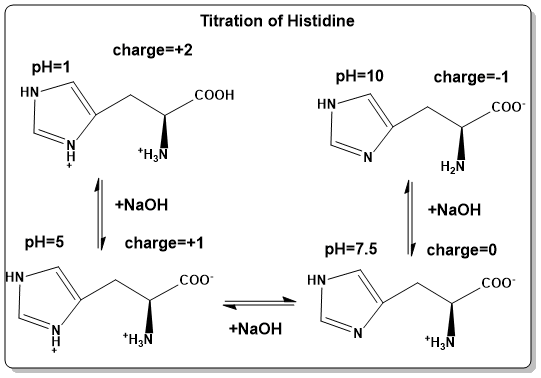
To illustrate the change of amino acid protonation with pH, histidine is shown as an example (Fig. 4). When dissolved in 0.1M HCl (pH=1) all ionisable groups are protonated. Hence, the carboxyl-group will be –COOH, the amino-group will be –NH3+ and one of the nitrogens in the imidazole ring will be protonated as well =NH+-. Due to the delocalisation of the positive charge in the imidazole ring, acidification does not protonate both nitrogens. To this solution 0.1M NaOH is added dropwise and the pH followed. At pH 5.0, the most acidic group (the α-carboxyl-group) will be fully dissociated to –COO–. Further titration will cause dissociation of the proton associated with the imidazole ring (=N-), which is largely accomplished at pH 7.5. Titration to pH 10, will see dissociation of the proton from the α-amino group (-NH2). The amino acid acts as a pH buffer at its pKa values. Let us use the pKa value of the α-carboxyl-group as an example. At pH 3.1, 50% of the α-carboxyl-group will be protonated (-COOH) and 50% will be deprotonated (-COO–). Addition of protons will cause some of the -COO– to become -COOH, thereby absorbing the protons that would make the solution more acidic. Vice versa adding OH– will cause some -COOH to lose a proton, which neutralises the OH- to form water. Once this process is exhausted the buffering power ceases.
Notably, the listed pK (Table 2) values apply to free amino acids in aqueous solution. At first glance only histidine appears to have a side-chain the charge of which can change in the physiological range of pH 6-8. However, the pK-value of amino acid side-chains can change dramatically depending on the environment. For instance, a more hydrophobic environment favours functional groups to be uncharged. Thus, a carboxyl-group is forced to remain protonated (-COOH). As an example, the side-chain carboxyl-group of glutamate, which in aqueous solution has a pK = 4.1, is often found protonated in the active site of enzymes, where it is surrounded by other amino acid side-chains and water. As a result, certain glutamate residues can act as an acid or as a base in biochemical reactions. Amino acid side-chains are often classified as polar when they have the ability to form hydrogen bonds or charge pairs, while being classified non-polar when they cannot form these interactions (aliphatic side chains). Non-polar (hydrophobic) amino acids are generally found in the core of proteins where they can interact with other hydrophobic side-chains instead of being exposed to polar water molecules. For the same reason non-polar amino acids are also enriched in transmembrane helices. The polar amino acid cysteine is special because the sulhydryl-groups (-SH) of two adjacent cysteine residues can oxidise to form a covalent disulfide bridge (-S-S-) (golden in Fig. 5). This bond is much stronger than any of the weak interactions formed by other amino acids. Disulfide bridges can only form in oxidising environments outside the cell, such as the blood, where you would find the hormone insulin (Fig. 5). We will revisit the properties of amino acids in more detail when looking at protein structure.

Test your knowledge
Q: What functional groups are common to all amino acids.
- Imidazole ring
- alpha-amino group
- delta-carboxyl group
- Guanidinium group
- alpha-carboxyl group
2 and 5, alpha-amino group and alpha-carboxyl group.
Nucleotides
Nucleotides are the building blocks of RNA and DNA molecules. However, nucleotides also serve important additional role as metabolites. The RNA building block ATP (Adenosinetriphosphate) is the major mobile energy carrier in all organisms. GTP (Guanosinetriphosphate), while used less universal, is used as an energy carrier in protein biosynthesis and in signal transduction processes. CTP (Cytidinetriphosphate) and UTP (Uridinetriphosphate) are used as energy carriers for carbohydrate conjugation. Nucleotides are a compound building block, made up of a carbohydrate (ribose or deoxyribose), a nucleobase (adenine (A), thymine (T), guanine (G), cytosine (C), and uracil (U)) and phosphate groups. In RNA ribonucleottides are used while in DNA deoxyribonucleotides are used.
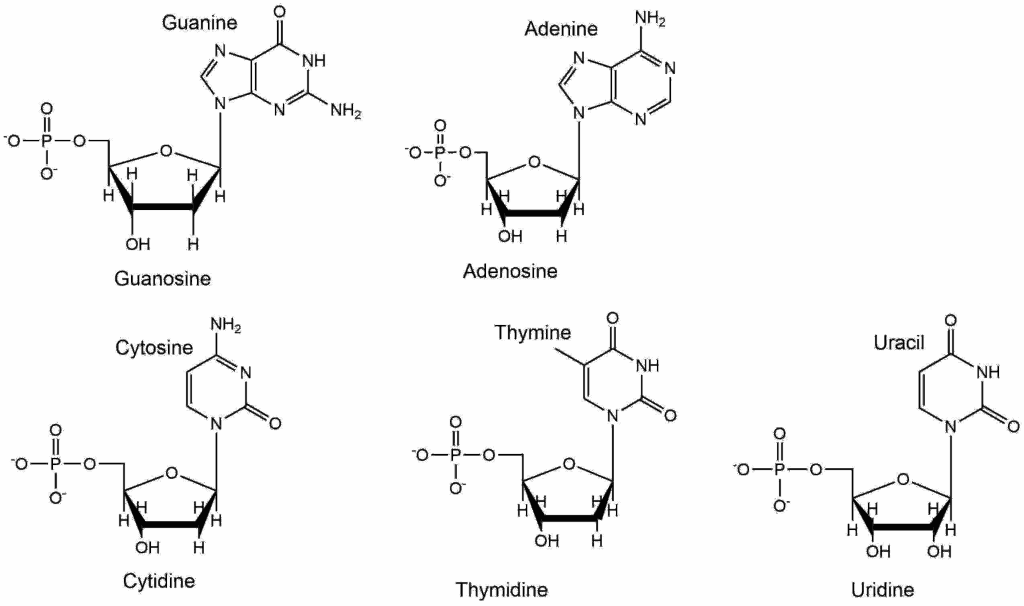
The naming convention is adenine, guanine, cytosine, thymine and uracil for the nucleobases. When attached to ribose they are called nucleosides and named adenosine, thymidine, guanosine, cytidine and uridine. They are called nucleotides when they contain phosphate groups in addition, which are attached to the C5 of ribose. To avoid confusion with the carbon atoms in the nucleobase the numbering is used with a prime e.g. 5’ or 3’.
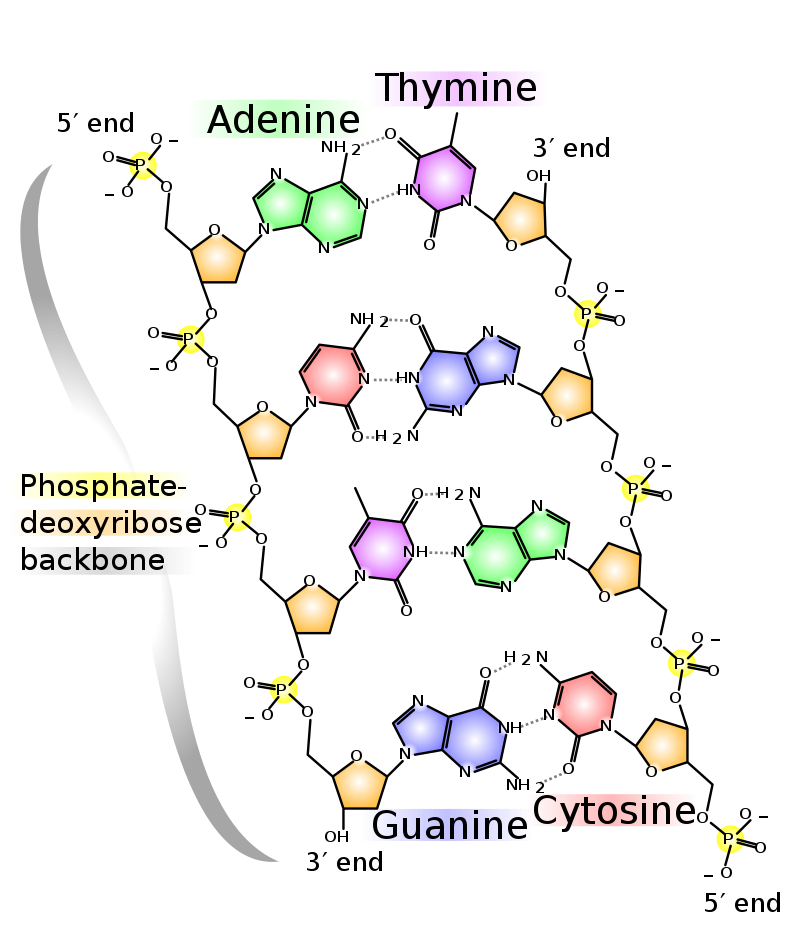
This can be used to indicate the direction of DNA polymerization. In DNA, 2-deoxyribose is used in nucleotides and A,T,G,C are used as nucleobases. In RNA ribose is used and A,U,G,C are the bases. The chemical nature of the nucleobases allows the formation of parallel hydrogen bonds with 2 bonds occurring between A and T and 3 bonds occurring between G and C. It is worth noting that nucleobases are planar, but there is a distinct angle in the bond to ribose, which allows adoption of the well-known helical structure by complementary strands of oligonucleotides. All cells can synthesize nucleotides de novo but are also able to salvage nucleotides from the circulation.
Lipids
Membranes enclose all cells and also their sub-compartments, such as lysosomes, peroxisomes, mitochondria, the endoplasmic reticulum and the Golgi apparatus. Vesicles are small membrane enclosed spheres that mediate the traffic of membrane lipids and of water-soluble contents between cellular compartments. The main feature of lipid membranes is their impermeability to water and water-soluble metabolites. Only through proteins, such as transporters or channels can ions and water-soluble metabolites cross the membrane. Biological membranes are comprised of a bilayer of lipids. This is a consequence of their physical properties. Similar to olive oil forming a droplet in water, membrane lipids form droplets in water due to their water repelling nature (also called hydrophobicity). It is energetically favourable for lipids to stick to each other and for water molecules to stick to each other (hydrophobic effect). In this arrangement water molecules can form hydrogen bonds with each other, and lipids can form weak bonds through van der Waals forces. In contrast to olive oil, membrane lipids also have a polar end, which is called the head-group (Fig. 8D: 1). As a result, membrane lipids are amphiphilic (having a polar and a non-polar side) because they interact with water through polar head-groups (see Fig. 8) and aggregate with their hydrophobic fatty acids (Fig. 8D: 2) to form the membrane. Some common lipids are shown in Figure 8 and 9.
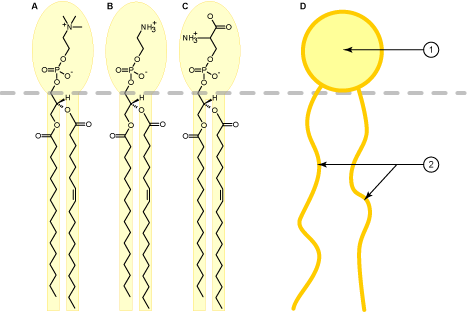
The amphiphilic structure of membrane lipids causes them to form bilayers, because it is energetically favourable for the hydrophobic parts to be in contact with each other as shown in the molecular dyanmics simulation video below. The video also demonstrates that a membrane is highly fluidic.

There are many types of lipids in cells. They are most readily grouped by the molecule to which fatty acids and polar groups can be attached (Fig. 9), namely glycerolipids (glycerol backbone), sphingolipids (sphingosine backbone) and cholesterol (sterol backbone).
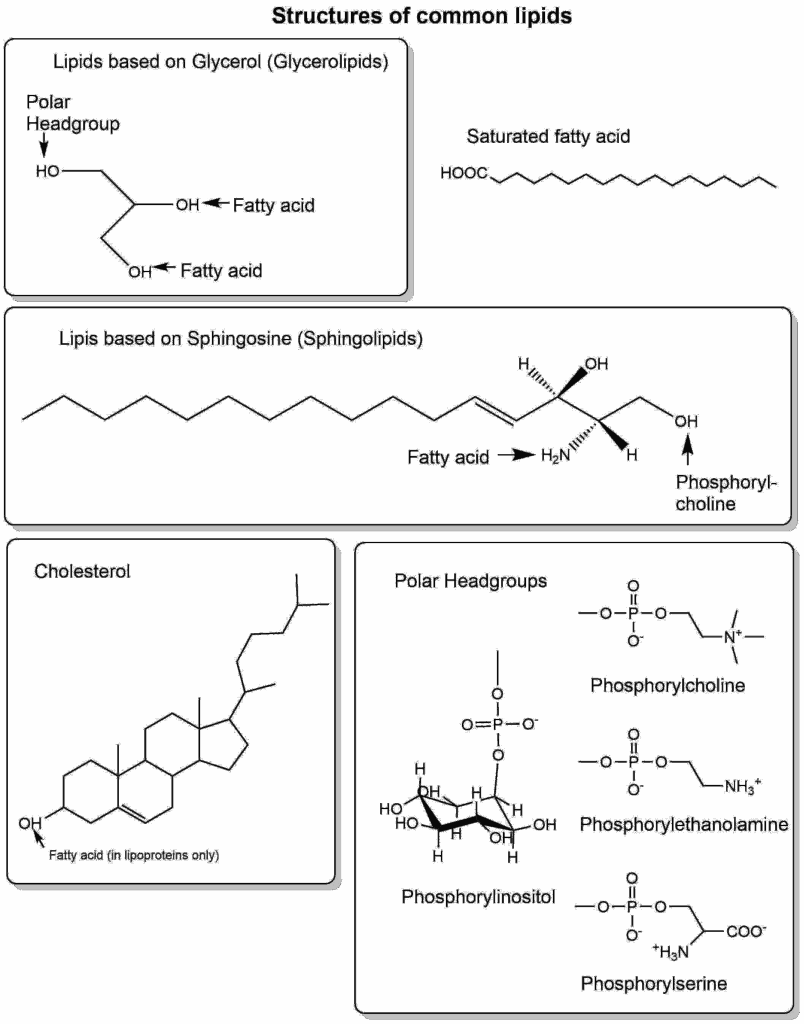
Glycerolipids can have a variety of head-groups, giving the lipid different biophysical and biochemical properties (Table 3).
| Headgroup | Structure | Property | Function |
|---|---|---|---|
|
Phosphoryl-ethanolamine |
-PO4-CH2-CH2-NH3+ |
Basic |
Membrane structure |
|
Phosphoryl-choline |
-PO4-CH2-CH2-N(CH3)3+ |
Basic |
Membrane structure |
|
Phosphoryl-glycerol |
-PO4-CH2-CH(OH)-CH2OH |
Neutral |
Membrane structure |
|
Phosphatidyl-inositol |
-PO4-CH6H11O6 |
Neutral |
Signalling |
|
Phosphoryl-serine |
H3N+-CH(CH2-PO4)-COO- |
Zwitterionic |
Signalling |
Carbohydrates
Carbohydrates are typically viewed as energy metabolites, but are also important building blocks. The surface of all cells is coated with complex carbohydrate-chains that are attached to proteins (N-glycosylation attached to glutamine and asparagine residues, O-glycosylation attached to serine, threonine and tyrosine residues) and glycolipids (sphingolipids with complex carbohydrate-chains). Carbohydrates are particularly prevalent in proteoglycans, which are extracellular proteins with large and complex carbohydrate-chains attached to them (Fig. 10).
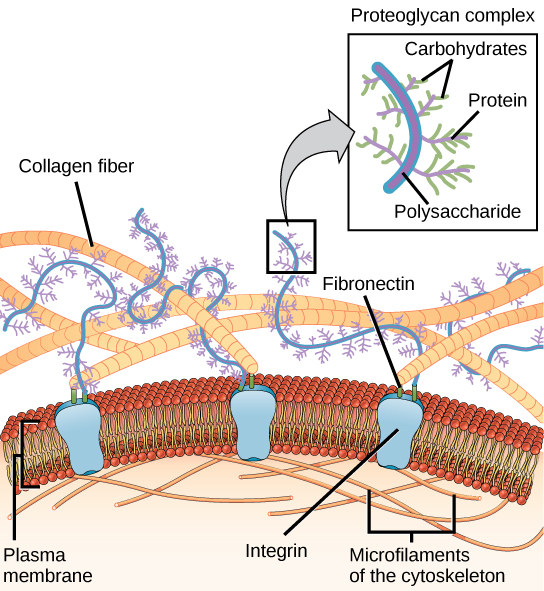
Proteoglycans have modified carbohydrates attached to them, containing amino groups, carboxyl-groups, sulfate groups etc. This allows holding more ions and water molecules within the extracellular space. This provides elasticity for example to our skin. You can squeeze your skin and feel the elasticity of the extracellular matrix. Proteoglycans are found in higher density in bone and cartilage. Another important role of carbohydrates is to provide visibility to the immune system. All extracellular proteins are coated with specific carbohydrates. Moreover, 2-Deoxyribose and Ribose form a building block in DNA and RNA, respectively.
Uric acid (Fig. 11) is often elevated in blood of people with metabolic syndrome (an early stage of type II diabetes). It has low solubility in water.
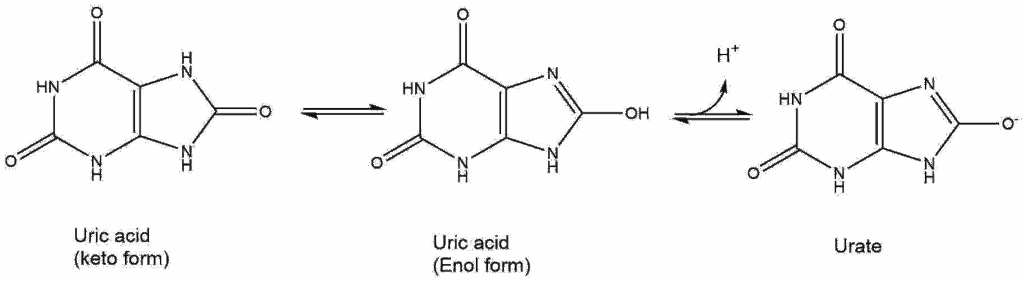
Answer the following questions:
Q1: Do you think it is a strong acid? Give a reason.
Q2: Compare uric acid to building blocks in the body, from what molecule might it be derived from?
Q3: What problem might the low solubility cause when there is too much uric acid (answer not covered in the text)?
It is not a strong acid. A strong acid is characterised by complete dissociation. The image shows two different tautomeres in equilibrium.Thus some of the acid is in the ketoform where it cannot dissociate.
Urica acid looks similar to nucleobases. Our body has no metabolic pathway to completely breakdown nucleobases. Excess nucleobases is released with the urine.
Low solubility can cause precipitates to form. This can occur in the kidneys generating kidney stones, but more typically in joints causing gout.
Q4. Match the following lipids to their class. Write down and toggle to class. Choices: Sterol lipid, Glycerolipid, Sphingolipid, Polar headgroup, Fatty acid.
- Phosphatidylserine
- Sphingomyelin
- Phosphorylcholine
- Palmitate
- Cholesterol
- Phosphatidylcholine
- Glycerolipid
- Sphingolipid
- Polar headgroup
- Fatty acid
- Sterol lipid
- Glycerolipid
Q5. Which structure of leucine is the most prevalent at pH 7.5?
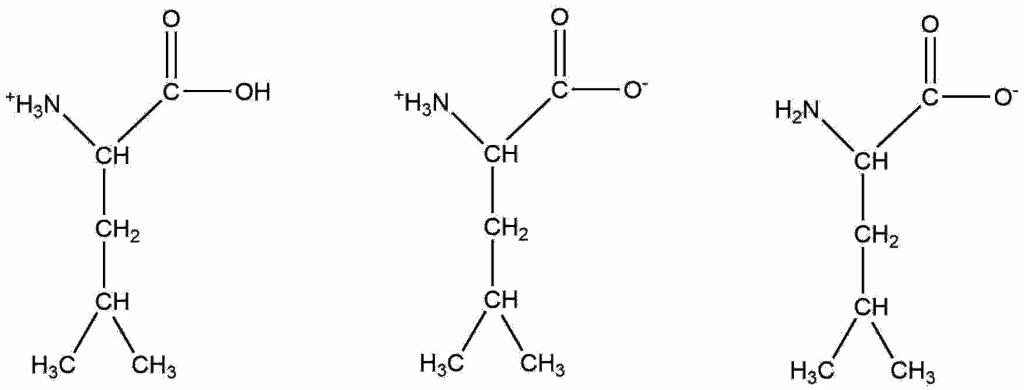
The structure in the center.
Q6. Assign each amino acid to the group with matching biophysical properties. (Choices: anionic, non-polar neutral, polar neutral, cationic, aromatic).
- Leucine
- Arginine
- Glutamate
- Phenylalanine
- Glutamine
- Non-polar neutral
- Cationic
- Anionic
- Aromatic
- Polar neutral
- Our major nutrients also form or are the major building blocks of a cell.
- Proteins are made up of 20 different proteinogenic L-amino acids
- Amino acids have an alpha-carboxyl group, an alpha-amino group and a side-chain. Free amino acids change charge at different pH, because of their acid/base properties.
- Amino acids, depending on their side chain, are grouped by their biophysical properties into neutral non-polar, neutral polar, neutral aromatic, negatively charged and positively charged amino acids.
- Nucleotides are a compound building block, which is made up of a carbohydrate (deoxy-ribose or ribose) a nucleobase and phosphate. Nucleobases can form hydrogen bonds with each other resulting in the well-known base pairing rules of DNA.
- With the exception of cholesterol, lipids are compound building blocks, which are made up of several smaller building blocks. They contain fatty acids, which form esters with their glycerol or sphingosine backbone. They also contain a polar head group which is linked to the backbone via a phosphodiether linkage. Cholesterol by contrast does not have different head-groups.
- Lipids are amphiphilic (water loving and oil loving). In membranes the polar head groups point towards aqueous compartments, while the fatty acids from many lipids align to form a bilayer. The bilayer is formed spontaneously, because it allows contact with water on both sides.
- Carbohydrates are important building blocks for nucleotides, they are attached to proteins and lipids.
- Proteoglycans (proteins with complex carbohydrate structures) are important for the structural integrity of tissue, skin, bone, cartilage, hair and nails.
- [3] By the author using ChemDraw
- [4] By the author using ChemDraw
- [5] By the author using PyMol
- [6] By the author using ChemDraw
- [9] By the author using ChemDraw
![[1] By Mediran – Own work, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=20664784](https://commons.wikimedia.org/w/index.php?search=by+mediran&title=Special%3ASearch&go=Go#/media/File:Eukaryotic_Cell_(animal).jpg){kind=link}
![[2] By TimVickers – Own work by uploader, based upon simlar illustrations in Goodsell DS (June 1991). “Inside a living cell”. Trends Biochem. Sci. 16 (6): 203–6. DOI:10.1016/0968-0004(91)90083-8. PMID 1891800., Public Domain, https://commons.wikimedia.org/w/index.php?curid=4762672](https://upload.wikimedia.org/wikipedia/commons/2/2e/Crowded_cytosol.png){kind=link}
![[7] By Madprime (talk · contribs) [CC0, GFDL (http://www.gnu.org/copyleft/fdl.html), CC-BY-SA-3.0 (http://creativecommons.org/licenses/by-sa/3.0/) or CC BY-SA 2.5 (https://creativecommons.org/licenses/by-sa/2.5)], from Wikimedia Commons](https://upload.wikimedia.org/wikipedia/commons/e/e4/DNA_chemical_structure.svg){kind=link}
![[8]By Foobar (self-made by Foobar) [GFDL (http://www.gnu.org/copyleft/fdl.html), CC-BY-SA-3.0 (http://creativecommons.org/licenses/by-sa/3.0/) or CC BY 2.5 (https://creativecommons.org/licenses/by/2.5)], via Wikimedia Commons](https://commons.wikimedia.org/wiki/File:Phospholipid_schematic_representation.png){kind=link}