
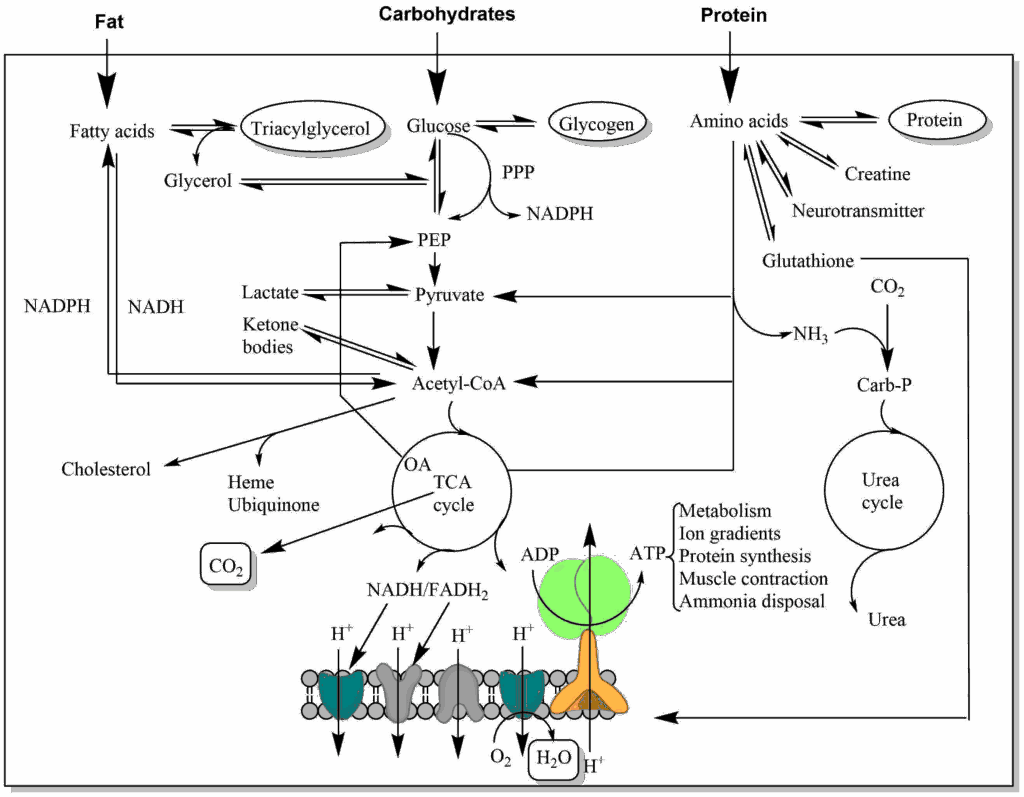
How do we know the energy content of different food groups? Wilbur Atwater and colleagues developed a room-sized human calorimeter for human nutrition studies in 1893 (see Image below). The exhaled air was passed through bottles of sulfuric acid to trap water and through bottle containing NaOH to trap carbon dioxide. Oxygen was provided from a cylinder. This allowed measuring the exchanges of water, carbon dioxide and oxygen by weighing the corresponding containers. In 1900 they published the caloric content of the major nutrients (in kcal/g) as 4, 4 and 9 for carbohydrates, protein and fat, respectively, which are still correct today.

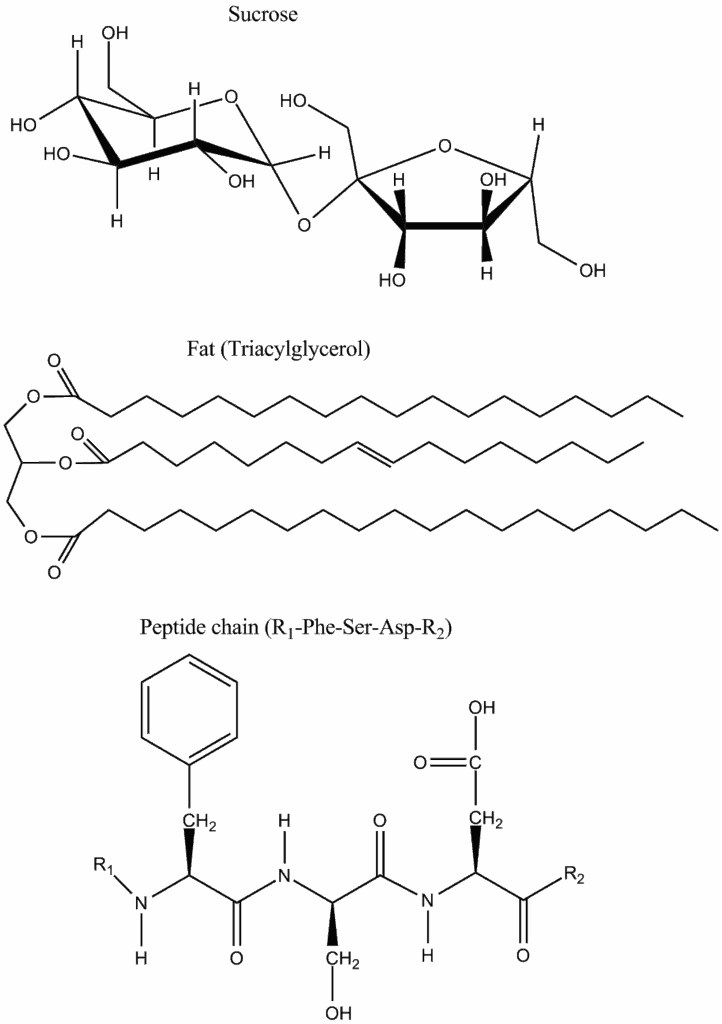
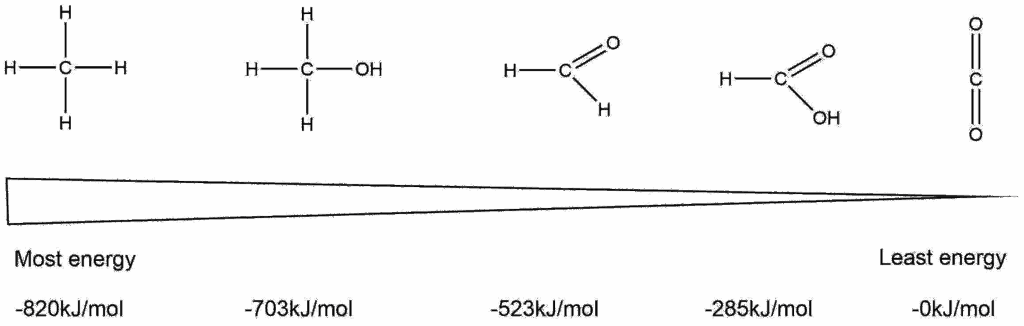
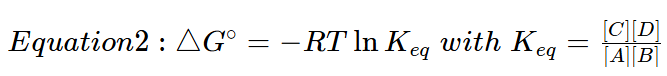
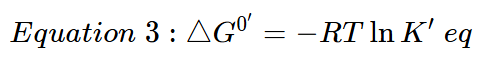
The impact of the concentration has an important consequence for biological reactions. All reactions involving water as a reactant (hydrolysis) will be driven by the high concentration of water in biological systems (55.5 M). Thus, hydrolysing a nutrient polymer such as starch or protein into its building blocks (sugar and amino acids) will always be favored and have a negative free energy (Table 2). Moreover, breaking a polymer down into individual building blocks also increases the entropy of the products.
To calculate the actual free energy using in vivo concentrations of reactants and products the following formula is used:
![]()
In this equation [A][B][C][D] are the prevailing in vivo concentrations, while the energy under standard conditions is incorporated into ΔG0′.
Full combustion of organic material is associated with a much larger negative free energy than hydrolysis, because we are starting with almost 100% reactants (for example cellulose) and end up with almost 100% products (CO2 and H2O). Please remember that the law of mass conservation applies. In the reaction C6H12O6 + 6O2 → 6CO2 + 6H2O the two substrates have the same weight as the two products. The products are much more stable than the substrates, because of a favourable electron configuration and because we are generating a large amount of gas molecules (H2O vapour and CO2), which have a far more random arrangement in space (higher entropy) than a solid block of wood.

![80 trading 24 [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], from Wikimedia Commons](https://commons.wikimedia.org/wiki/File:Bush_fire_at_Captain_Creek_central_Queensland_Australia..JPG){kind=link}
{kind=link}